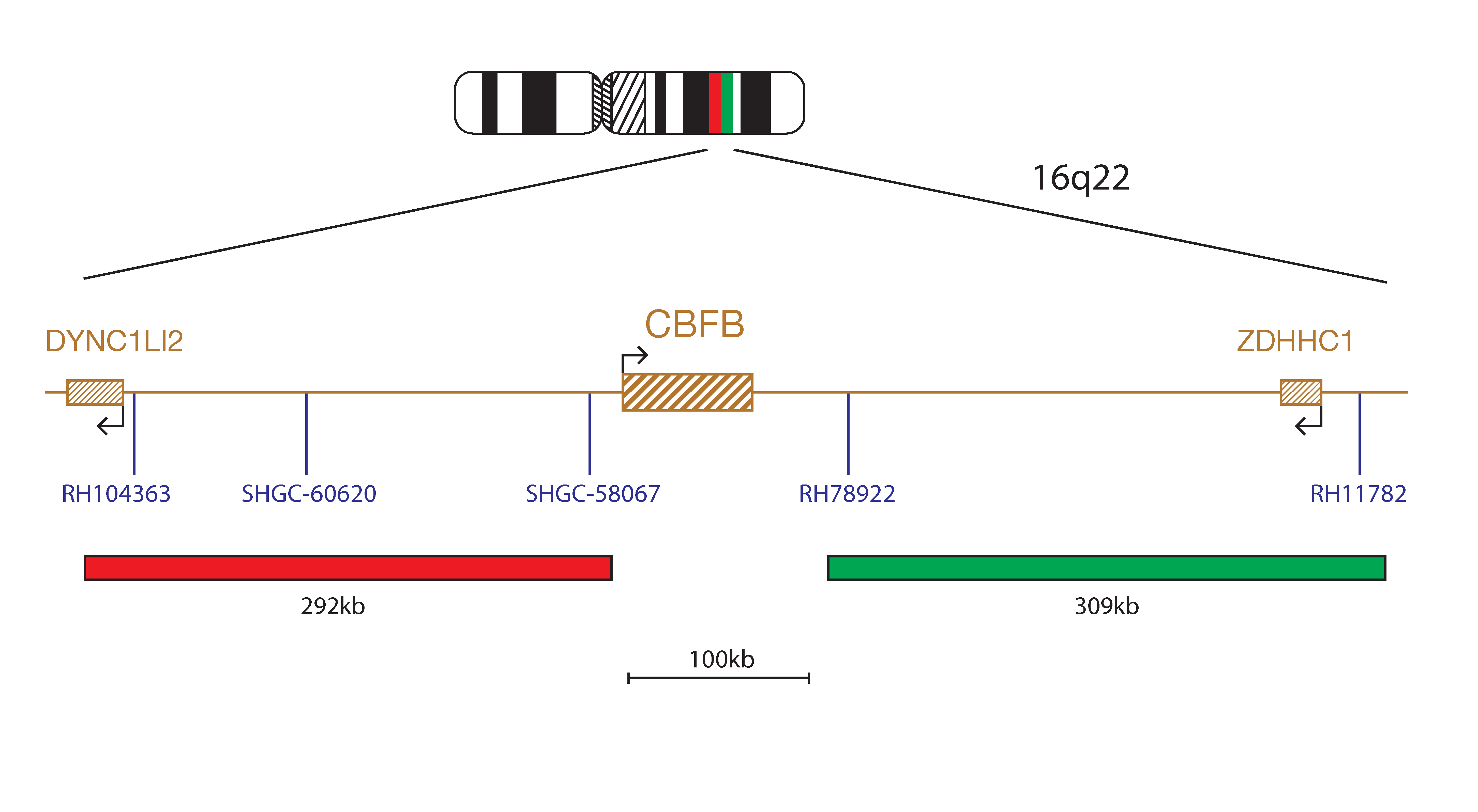
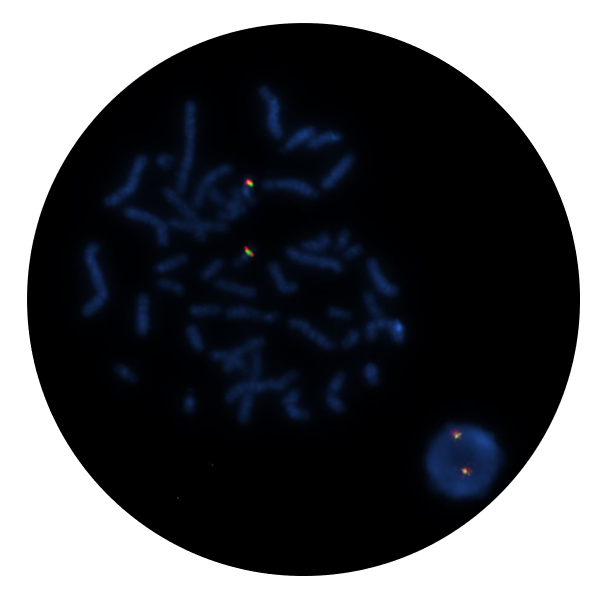
The CBFB Breakapart Probe mix consists of two distinct probes. The red probe (292kb) is centromeric to the CBFB gene and extends beyond the RH104363 marker to cover part of the DYNC1LI2 gene and includes the markers SHGC-60620 and SHGC-58067. The green probe (309kb) is telomeric to the CBFB gene and extends through the marker RH78922 beyond the ZDHHC1 gene to a region telomeric to the marker RH11782.
The CBFB (core-binding factor subunit beta) gene is located at 16q22, it is most commonly rearranged due to the inversion inv(16)(p13.11q22.1) or the translocation t(16;16)(p13.11;q22.1). Rarely, translocations of 16q22 with various other gene partners have been reported, whilst deletion of the band 16q22 has also been reported1.
Acute myeloid leukaemias with inv(16)(p13.11q22.1) or t(16;16)(p13.11;q22.1) form a recognised disease entity according to the World Health Organisation (WHO) classification of myeloid neoplasms and acute leukaemia2. These rearrangements are frequently found in patients with a myelomonocytic subtype with increased bone marrow eosinophils, formerly referred to as AML FAB (French-American-British classification) type M4Eo, and are found in 5-8%2 of AMLs. Cases of therapy-related AML may also have this rearrangement2,3.
Inversion inv(16)(p13.11q22.1) or translocation t(16;16)(p13.11;q22.1) produce CBFB-MYH11 gene rearrangements, and are classed as a favourable cytogenetic risk group in patients with AML4,5,6.
For both inversion inv(16)(p13.11q22.1) or the translocation t(16;16)(p13.11;q22.1), the breakpoints occur in intron 5 of CBFB and intron 5 of MYH11. The N-terminal of CBFB fuses to the C-terminal of MYH11 with its multimerization domain. The resultant chimaeric protein reduces the amount of active CBF. An accumulation of CBFB-MYH11/CBFA multimers in the nucleus also occurs. CBFB regulates the expression of certain ADP-ribosylation factors (AFRs) and other tumour suppressor genes (TSGs) and therefore the fusion protein is thought to repress TSG expression4. This fusion protein has been shown to be necessary, but not solely sufficient for leukaemogenesis, and work continues to determine how the protein appears to work collaboratively with RUNX1 to mediate the proliferative signal and cell differentiation block required for development of leukaemia7,8.
I first came across CytoCell FISH probes in a previous lab I worked in and I was struck by the quality of the products. Since this time, I have been recommending and introducing CytoCell probes across all application areas — now they are the primary FISH probes used in our lab. They have an excellent range of products and their ready-to-use reagent format saves considerable time.

Elizabeth Benner
Medical Technologist, University of Arizona Health Network, USA

 Visit USA site
Visit USA site Visit Canada site
Visit Canada site